Впервые с использование микросателлитных маркеров оценено генетическое разнообразие сибирской косули (Capreolus pygargus). Установлено наличие генетических различий между западными (Урал и Западная Сибирь) и восточными (Забайкалье и Дальний Восток) группировками вида, которые, возможно, отражают географию размещения подвидов
В настоящее время генетика европейской косули (Capreolus capreolus) исследована довольно хорошо, в то время как данные для сибирской косули (C. pygargus) весьма ограничены. Совместно с коллегами из Сеульского национального университета, Национального университета Монголии, Лазовского заповедника и Института биологических проблем криолитозоны СО РАН изучены генетические характеристики, структура населения и связи между географически разобщенными группировками вида по 12 микросателлитным локусам (табл.).
Таблица – Показатели генетического разнообразия в исследованных выборках
|
Регион |
N |
СЧА |
Ar |
He |
Ho |
ЧСА (диапазон частот) |
|
SKJ |
33 |
3.75 |
2.18 |
0.386 |
0.329 |
4 (0.016-0.106) |
|
SKM |
31 |
6.58 |
3.48 |
0.596 |
0.451 |
3 (0.016-0.065) |
|
RPR |
30 |
7.42 |
3.67 |
0.623 |
0.490 |
4 (0.017-0.050) |
|
RSMG |
21 |
7.00 |
5.67 |
0.598 |
0.500 |
7 (0.024-0.025) |
|
RYA |
18 |
5.33 |
3.26 |
0.553 |
0.459 |
5 (0.031-0.094) |
|
RARN |
12 |
3.92 |
3.87 |
0.560 |
0.503 |
0 |
|
RURK |
44 |
4.92 |
3.73 |
0.534 |
0.495 |
3 (0.011-0.012) |
Для всех группировок характерен средний уровень генетического разнообразия (Не варьировал от 0.522 до 0.628); для популяции на острове Чжеджу (Южная Корея) этот показатель был заметно ниже (Не=0.386). Для группировок из западной части ареала (Урал и Западная Сибирь) характерны более низкие показатели аллельного разнообразия по сравнению с группировками из восточной части ареала (Южная Сибирь, Забайкалье, Приморье, Корейский полуостров): среднее значение аллельного разнообразия (Ar) для западной части ареала составило 2.81, для восточной – 3.54.
Кластерный анализ генетических дистанций Da позволил разделить рассмотренные выборки на три основные клады (рис .1). В первую вошли выборки из западной части ареала (Урал и Западная Сибирь), во вторую – из восточной и центральной частей ареала (Забайкалье, Приморье, Корея). Выборка из Алтайского края занимает промежуточное положение. Причину выделения этой популяции трудно оценить из-за малого объема выборки (n=5). Изолированная группировка (остров Чеджу) образовывает отдельную кладу. Кластерный анализ, выполненный с помощью программы K-structure, также подтверждает выделение указанных клад и подчеркивает смешанный характер выборки из Якутии. Внутри «западной» и «восточной» группировок генетической дифференциации практически не выявлено.
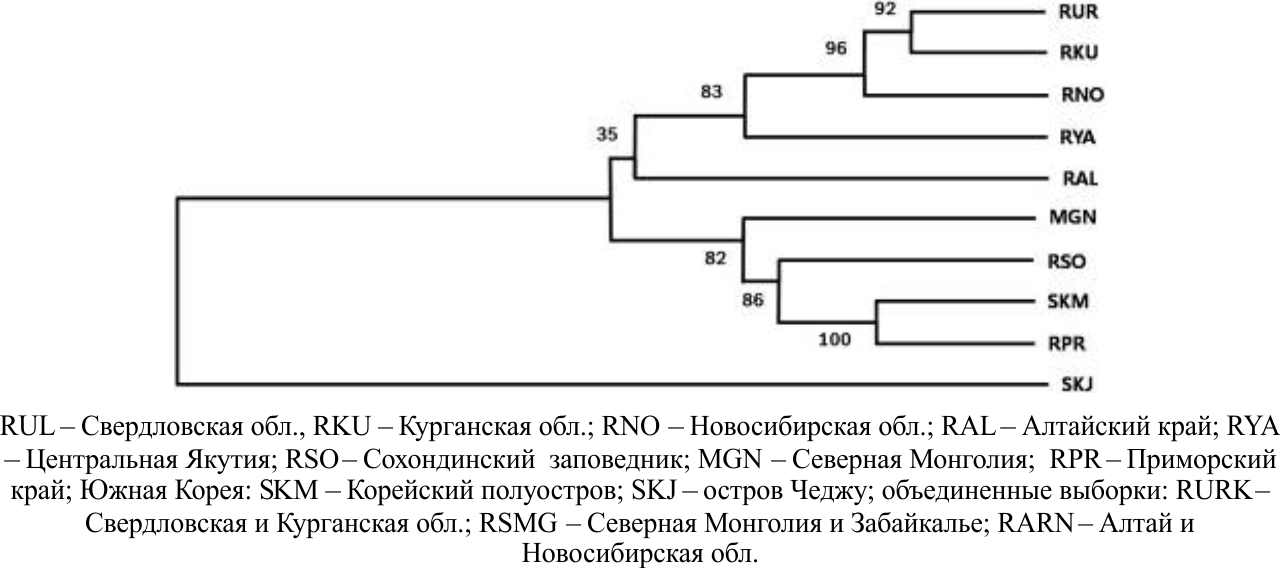
Рисунок 1 – Дендрограмма связей между рассмотренными выборками по генетическим дистанциям
Генетические дистанции между рассмотренными группировками на 37,6% определись географическими дистанциями. Наиболее сильное влияние расстояние между районами сбора данных оказывало на различия между «смешанными» (Алтай, Монголия) и «западными» выборками. В то же время более 60% различий не могут быть объяснены пространственной разобщенностью выборок, по-видимому, большее значение в их генетической дифференциации имеют другие факторы, такие как изоляция, возможно, и отбор.
Для анализа роли пространственной изоляции в формировании наблюдаемых генетических группировок сибирской косули выдвинута гипотеза о роли горных хребтов, как фактора их генетической дифференциации. Анализ, основанный на корреляции генетических и географических дистанций, позволил выявить зоны «перепадов» этих параметров. На рис. 2 показано размещение этих «барьеров», которые в целом совпадают с размещением основных горных хребтов в Южной Сибири, а также с границами ранее описанных подвидов сибирской косули Caprolus pygargus pygargus и C. p. tianschanicus (Гептнер и др., 1961). Таким образом, горные хребты Южной Сибири могли выступать в качестве природного фактора, ограничивающего обмен генами между животными в юго-западной части ареала. С другой стороны, Становой хребет не служит границей распространения «восточной» группы к северу, и генетический состав якутской выборки свидетельствует о возможной гибридизации представителей западной и восточной групп на северной периферии ареала. Полученные результаты свидетельствуют об особом статусе популяции острова Чеджу, который является результатом долговременной изоляции и адаптации к условиям обитания на данном острове. Дальнейшие исследования позволят уточнить таксономический статус этой группировки.
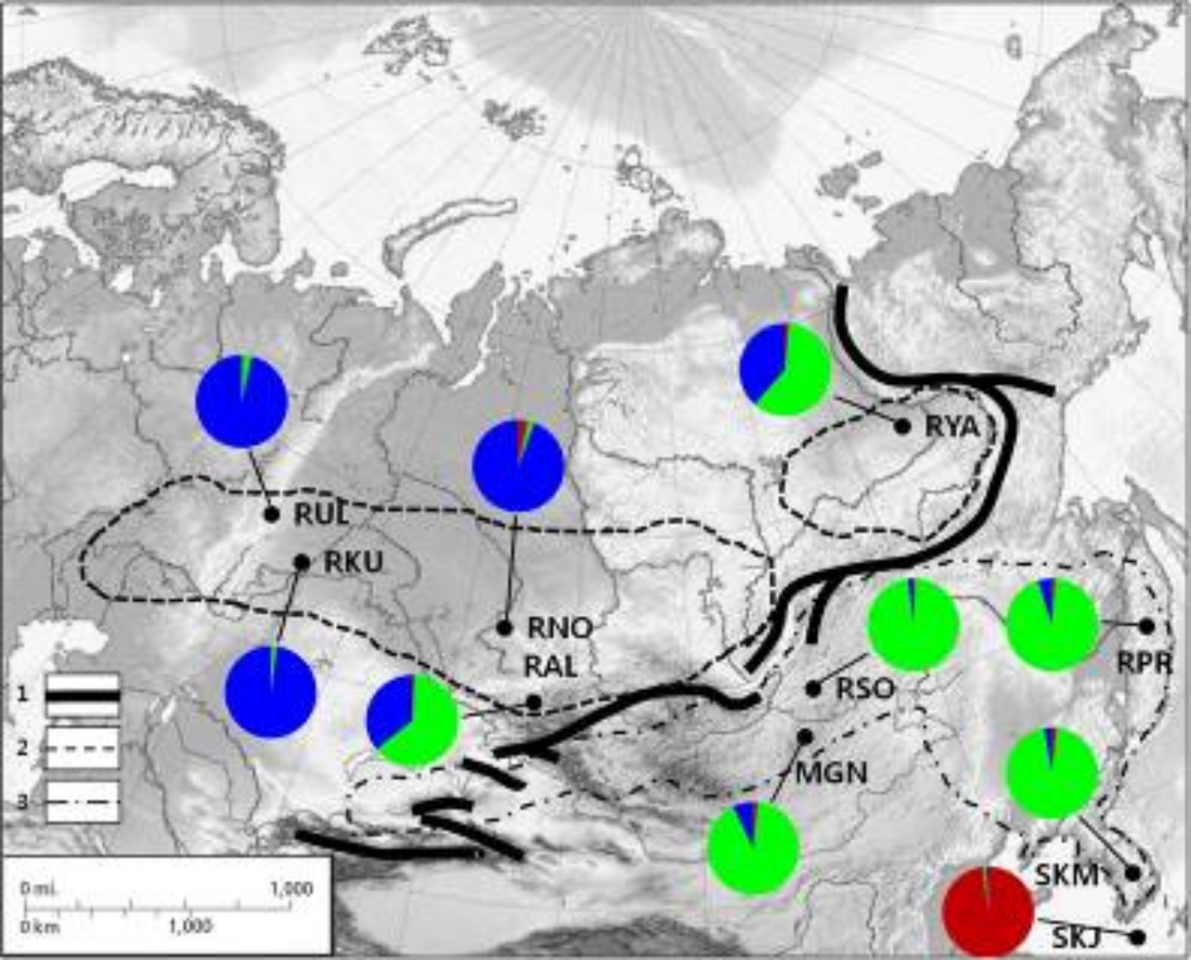
Рисунок 2 – Географическое размещение аллельных групп в исследованных выборках сибирской косули: 1 – горные хребты; границы ареала (по Данилкину, 1999) подвидов C.p.pygargus (2) и C.p.tianschanicus (3)