Выявлены закономерности географической изменчивости размеров имаго белянок (Lepidoptera: Pieridae: Pierini) Уральского региона, построены карты вольтинности основных видов. Впервые для анализа сезонной фенотипической пластичности насекомых применен метод морфофизиологических индикаторов. Показано, что сезонные генерации брюквенницы различаются по относительным размерам тех признаков, которые связаны с плодовитостью и приспособленностью имаго к расселению
С целью прогнозирования числа вылетающих сезонных поколений бабочек-белянок рода Pieris, являющихся вредителями сельского хозяйства, с использованием ГИС-технологий построены карты проявления вольтинности (числа поколений) видов в разных биомах Уральского региона (рис. 1). Для моделирования были использованы усредненные многолетние климатические данные. В северных районах реальное число поколений может превышать прогнозируемое, поскольку не учитываются кратковременные потепления и дневные нагревы, достаточные для развития. Следует также учитывать, что важную роль в распространении белянок могут также играть трофические связи с кормовыми растениями. В целом, полученная карта хорошо согласуется с литературными данными и нашими наблюдениями.
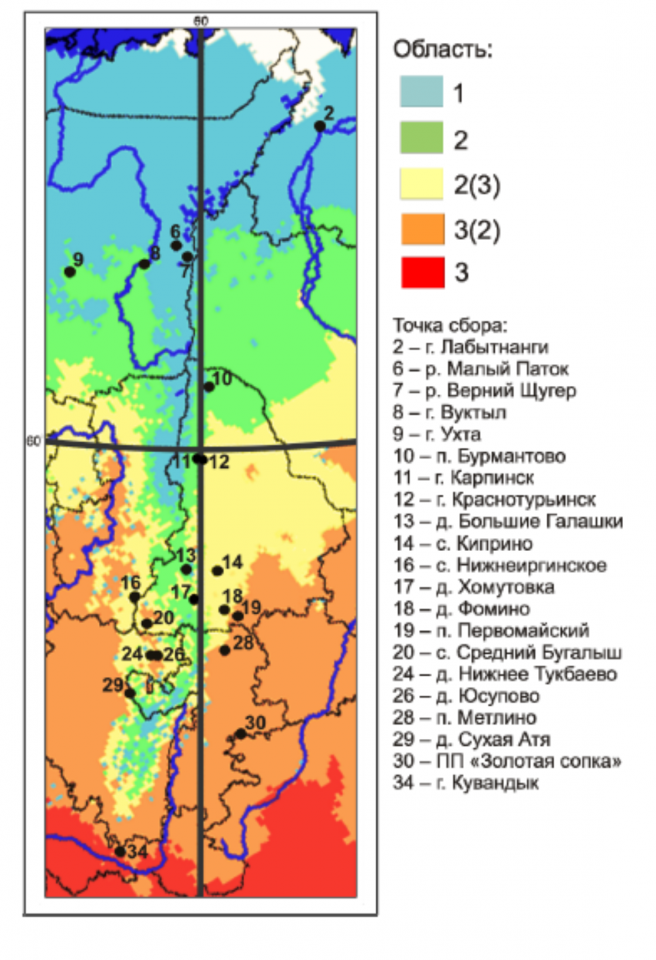
Рисунок 1 – Районирование Уральского региона по количеству генераций имаго репницы (Pieris rapae) за теплый сезон
Анализ географической изменчивости размеров крыльев имаго первой генерации P. napi позволил установить, что имаго из области, где для данного вида характерен моновольтинный жизненный цикл, значимо крупнее, чем имаго из других областей Уральского региона. Сравнение площади крыльев имаго второй генерации P. napi показало, что в области, где второе поколение факультативно, бабочки обладают наименьшими по размеру крыльями (рис. 2).
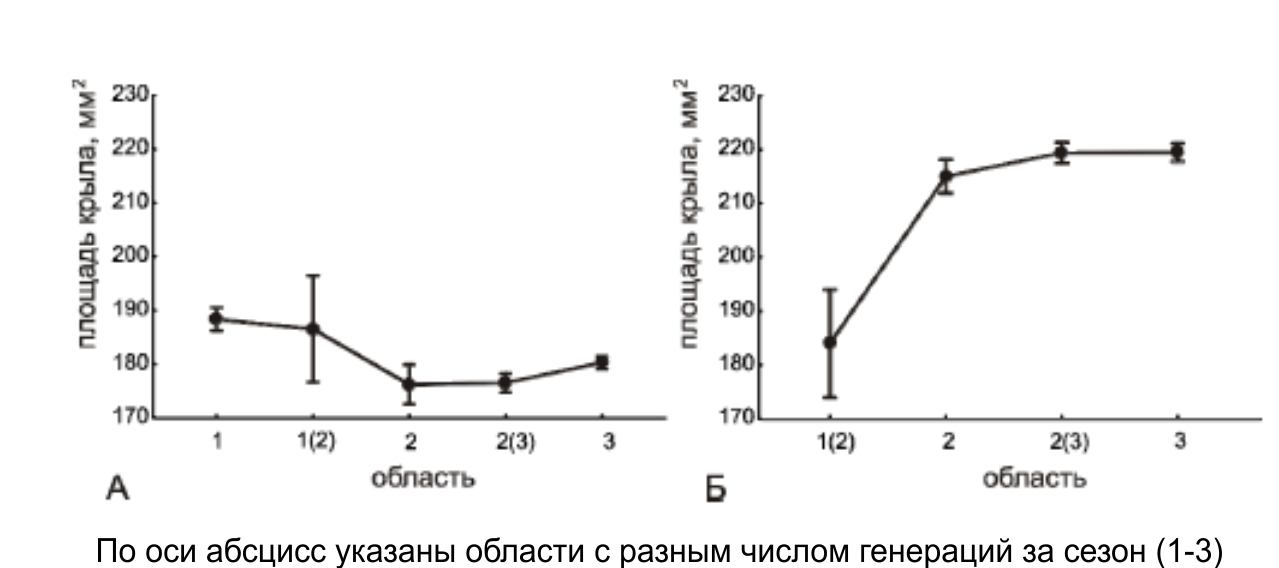
Рисунок 2 – Географическая изменчивость размеров крыла первой (А) и второй (Б) генераций имаго P. napi
Проявления географической изменчивости различны у разных генераций имаго. В пределах весенней генерации прослеживается обратная зависимость между площадью крыла и количеством генераций в исследуемой географической области. Вероятно, это связано с приспособлением географических популяций к различным климатическим условиям. В северных широтах и горных районах продолжительность и средняя температура благоприятного для развития периода меньше, также возможно снижение температуры ниже пороговой для данных видов. В таких условиях более выгоден медленный и продолжительный рост, при котором не снижается устойчивость к различным стрессовым факторам, в результате имаго достигают более крупных размеров. Если условия допускают развитие факультативной генерации, то для недиапаузирующей части популяции уменьшение продолжительности роста гусениц оказывается выгодным, поскольку необходимо завершить развитие в более сжатые сроки до наступления неблагоприятного периода. Сокращение периода развития преимагинальных стадий приводит к уменьшению размеров имаго факультативной генерации.
В пределах Уральского региона существует фенологический сдвиг развития сезонных генераций при поливольтинном жизненном цикле. Так, условия развития второй генерации из более северного района сходны с условиями развития третьей генерации из более южного. Кроме того, развитие генерации при моновольтинном жизненном цикле проходит в летние месяцы, т.е. при относительно длинном световом дне. Чем больше генераций наблюдается за сезон, тем на более поздние сроки сдвигается развитие первой, диапаузирующей, генерации и, соответственно, короче световой день, при котором она развивается.
Проведен анализ фенотипической пластичности морфофизиологических признаков, связанных с плодовитостью и способностью к расселению, у поливольтинного вида P. napi на территории Южного Урала (окрестности п. Метлино Каслинского р-на Челябинской обл.). Статистически значимая регрессионная зависимость массы брюшка имаго от дня отлова установлена для обоих полов: в течение лёта первой генерации средняя величина массы брюшка снижается (рис. 3 А). Для обоих полов установлено статистически значимое уменьшение среднего значения относительной массы брюшка в популяции к концу лёта первого поколения (рис. 3 Б).
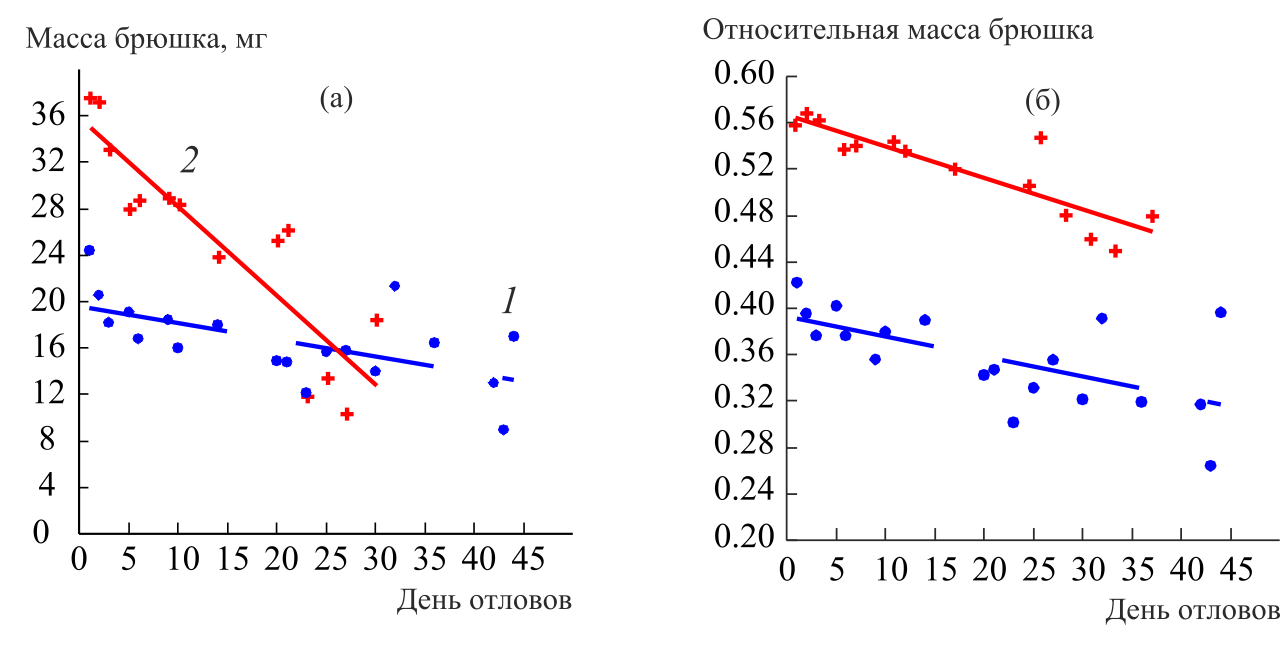
Рисунок 3 – Динамика абсолютной (а) и относительной (б) массы брюшка имаго самцов (1) и самок (2) P. napi в течение лёта весенней генерации 2011 г.
С помощью однофакторного дисперсионного анализа проведена оценка сезонной изменчивости относительной массы брюшка самцов и самок P. napi. В течение лёта одной генерации относительная масса брюшка уменьшается, а с массовым вылетом имаго следующей генерации вновь возрастает (рис. 4). Наибольшей относительной массой брюшка обладают особи весеннего поколения. При анализе сезонной динамики относительной массы брюшка можно обнаружить начало вылета осенней генерации P. napi (от периода 4 к периоду 5), чего не удается сделать по абсолютным показателям массы тела или другим признакам.
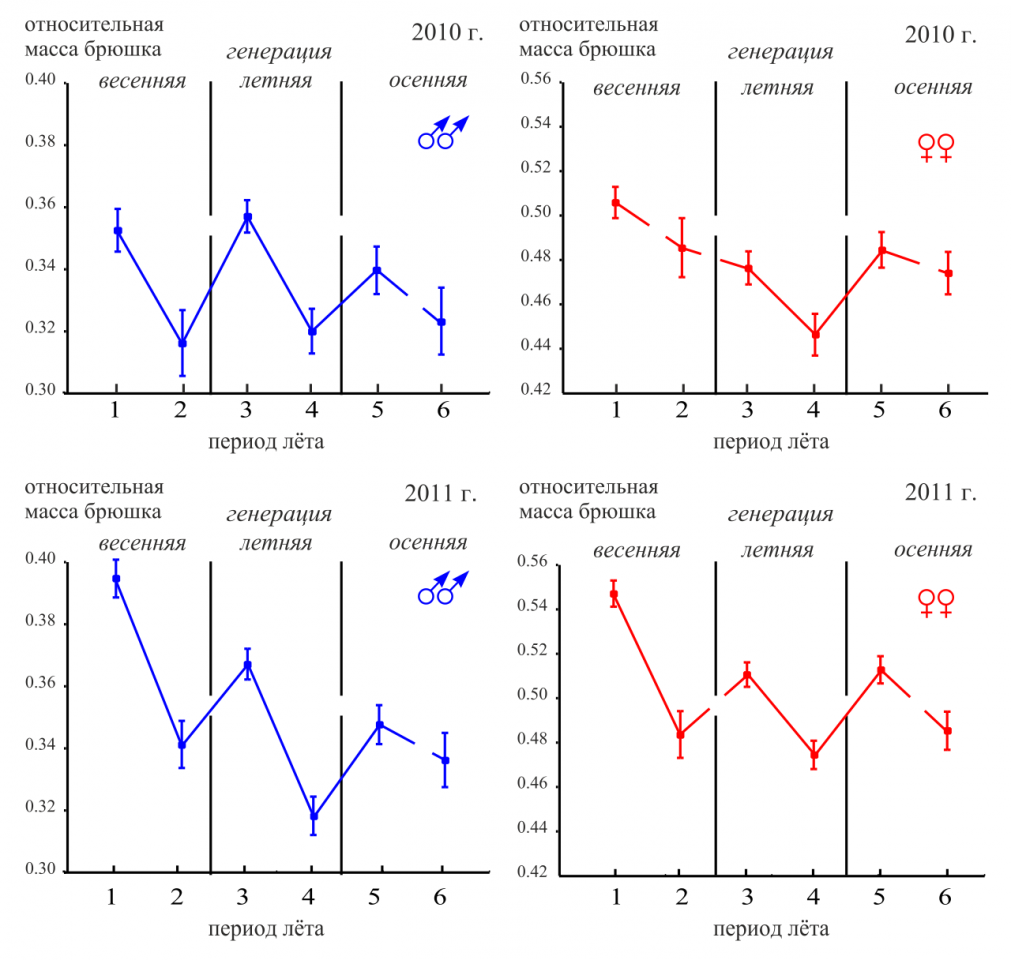
Рисунок 4 – Сезонная изменчивость относительной массы брюшка самцов (а) и самок (б) P. napi в 2010-2011 гг.